







Los viñedos enfrentan numerosas plagas y enfermedades, como por ejemplo la dañina lobesia botrana o polilla de la vid, y para combatirlos dependen en gran medida de insecticidas sintéticos, los cuales también afectan a organismos que no son objetivo de esos agroquímicos. En las viñas orgánicas y en los cultivos con mayor biodiversidad y mayor cobertura vegetal se dan las condiciones para que aves, murciélagos y artrópodos depredadores contribuyan a controlar esas plagas. En el estudio que aquí replicamos, un grupo de científicos húngaros e italianos investigaron el papel de estos animales en el control biológico de plagas en paisajes con amplia cobertura forestal.
Los viñedos son sistemas culturales, ecológicos y económicos de gran relevancia, que cubren aproximadamente 7,2 millones de hectáreas en todo el mundo, de las cuales alrededor del 50% se concentran en Europa (OIV, 2023). Se espera que esta superficie se expanda debido a la alta demanda de servicios de aprovisionamiento y culturales relacionados, así como a la creciente disponibilidad de áreas explotables en los límites más fríos del rango potencial de la vid como consecuencia del cambio climático.
Los viñedos enfrentan numerosas plagas y enfermedades y, a pesar de las tendencias de mejora, sólo áreas relativamente pequeñas adoptan prácticas de manejo sostenible (por ejemplo, solo el 12% de los viñedos europeos son orgánicos). La mayoría de los viñedos aún depende en gran medida de los tratamientos con pesticidas, los cuales afectan negativamente a organismos no objetivo.
Por ejemplo, la polilla del racimo de la vid (Lobesia botrana), una plaga ampliamente distribuida y significativa que causa daños severos y pérdidas de rendimiento, se controla principalmente con insecticidas sintéticos, lo que limita la regulación por enemigos naturales. Sin embargo, en condiciones adecuadas, los viñedos tienen el potencial de ofrecer servicios efectivos de control de plagas al fomentar comunidades de depredadores diversas y abundantes, como aves, murciélagos y artrópodos.
El control natural de plagas, proporcionado por una amplia gama de taxones vertebrados e invertebrados, es uno de los servicios ecosistémicos más importantes en los agrosistemas, con un valor económico estimado de 417 U$S por hectárea y por año en distintos biomas.
Entre los diversos grupos taxonómicos, las aves y los murciélagos son componentes clave de los servicios ecosistémicos basados en la biodiversidad y se consideran importantes agentes de supresión de plagas. Por ejemplo, las aves de zonas agrícolas consumen alrededor de 28 millones de toneladas de presas al año, incluidas grandes cantidades de insectos herbívoros, y los murciélagos consumen más del 70% de su masa corporal en artrópodos cada noche, con más de 700 especies de plagas identificadas en su dieta.
Al mismo tiempo, particularmente las aves pueden contribuir a ciertos diservicios, como la disminución de la intensidad de las cascadas tróficas al alimentarse de artrópodos depredadores.
La efectividad de estos servicios (o diservicios) depende en gran medida de factores que actúan a diferentes escalas espaciales. A diferencia de las prácticas convencionales de protección de cultivos, la agricultura orgánica, al incrementar la disponibilidad de recursos alimenticios asociada con la reducción en el uso de pesticidas, promueve un amplio espectro de funciones ecológicas y una alta proporción de especies de vertebrados con importancia en el control de plagas.
Otro factor influyente es el contexto del paisaje, que podría tener mayor relevancia que las características locales de las áreas agrícolas por sí solas. Por ejemplo, el incremento y la conectividad de los hábitats naturales no cultivados en el paisaje circundante son fundamentales para la biodiversidad de los viñedos, afectan positivamente la composición y actividad de las comunidades de murciélagos y favorecen una mayor riqueza y densidad de aves insectívoras.
La exclusión experimental constituye una herramienta poderosa para identificar interacciones complejas entre vertebrados e invertebrados que afectan los servicios ecosistémicos y el rendimiento de los cultivos. Sin embargo, los resultados previos se han reportado principalmente en áreas tropicales, y estudios de este tipo en agrosistemas permanentes europeos siguen siendo escasos.
Aunque las aves y los murciélagos suelen ser objeto de medidas de conservación, algunos agricultores aún tienden a invertir menos esfuerzo en atraerlos, pasando por alto los servicios de control de plagas que estos animales pueden proporcionar.
En este estudio, se realizó un experimento de exclusión en viñedos húngaros con prácticas de manejo y características de paisaje distintas, con el fin de investigar el papel de las aves y los murciélagos en la configuración de los patrones de densidad de artrópodos y en las funciones ecosistémicas asociadas. Se plantearon las siguientes hipótesis: (1) el manejo orgánico de los viñedos y los paisajes forestales aumentan la abundancia y actividad de aves y murciélagos; (2) un mayor número y actividad de aves y murciélagos resultan en una menor abundancia de insectos plaga; y además, (3) una reducción de la herbivoría de insectos y un aumento del rendimiento del cultivo, pero una menor presión de depredación por parte de los artrópodos.
Materiales y métodos: área y diseño del estudio
Este estudio se realizó en la parte occidental de las Montañas de Transdanubia (Cuenca de Tapolca y Tierras Altas de Balaton) y en la parte sur de la Pequeña Llanura Húngara (Cuenca de Marcal), Hungría (Figura S1). Esta región se caracteriza por un clima continental con influencia submediterránea, una temperatura media anual de 9–11 °C y una precipitación de 550–650 mm.
Para el experimento se seleccionaron 12 viñedos (que incluían 10 variedades de uva; una o dos variedades por plantación), 6 con manejo orgánico y otros 6 con manejo integrado de plagas (IPM). A diferencia de los viñedos IPM, los viñedos orgánicos (3,2% de los viñedos húngaros en 2025) se manejaron sin herbicidas, insecticidas sintéticos ni fertilizantes, y en nuestra zona de estudio presentaban generalmente menor superficie y rendimiento, pero mayor cobertura de vegetación herbácea (Tabla S1). En los viñedos IPM, el uso de insumos químicos se aplicó de manera limitada y selectiva en función del monitoreo de plagas, con el fin de mantener sus poblaciones por debajo de los niveles de daño al cultivo.
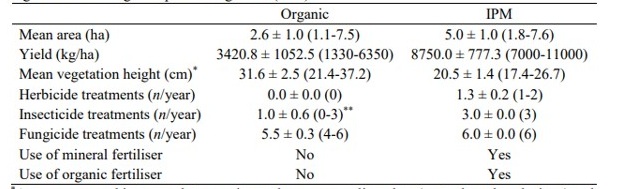
Los paisajes vitícolas se clasificaron como «forestados» y «no forestados» según su composición y configuración dentro de radios de 500 y 1000 m, basados en Corine Land Cover (Copernicus Land Monitoring Service, 2018), el Mapa de Ecosistemas de Hungría (Tanács et al., 2021) y en imágenes satelitales de Google (tomadas en 2023), utilizando el software QGIS 3.6.1 (QGIS Development Team, 2019) y Conefor Sensinode 2.6 (Saura & Torné, 2009).
En ambas escalas espaciales, los paisajes forestados tenían la mayor proporción de bosques caducifolios y la mayor conectividad de elementos leñosos (es decir, parches de bosque, setos, grandes grupos de árboles), mientras que los paisajes no forestados tenían una proporción relativamente alta de tierras de cultivo y las mayores distancias entre viñedos y bosques (Tabla S2). En total, se contó con tres réplicas de viñedos para cada combinación de factores (es decir, orgánico y forestado, orgánico y no forestado, IPM y forestado, IPM y no forestado). La distancia entre viñedos fue de entre 1 y 38 km (media ± EE: 16,7 ± 1,7).
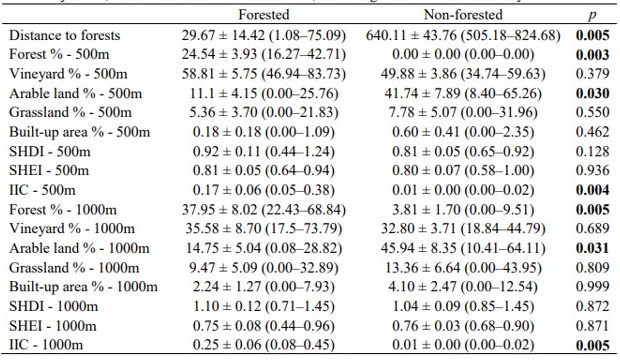
Se establecieron dos pares de parcelas control y exclusión completa (n = 24; en total 48 parcelas) en el centro de cada viñedo, con al menos 100 m de separación entre ellas y 25 m del borde. Para cada par, las parcelas control y exclusión estaban separadas por 25 m (Figura 1a).
Las jaulas de exclusión consistieron en cables de alambre rectangulares unidos con postes de bambú y cubiertos completamente con una malla agrícola desmontable (tamaño de malla de 2,5 × 2,5 cm) que permitía el acceso de insectos (incluidas polillas) a las plantas, pero impedía que aves y murciélagos alcanzaran el dosel. Cada jaula de exclusión medía 6 × 0,7 × 2 m (largo × ancho × alto) y siempre cubría seis plantas de vid (Figura S2). Se instalaron las jaulas en la primera quincena de abril de 2023, las retiramos a finales de agosto de 2023 antes de la cosecha y las mantuvimos regularmente durante el período de estudio. Dentro de cada viñedo, las vides control y excluidas se cultivaron de manera similar (por ejemplo, siega, poda, guiado de brotes y despunte).
Recolección de datos
Se utilizó el método de conteo por puntos (5 min por punto de observación, r = 50 m) para las observaciones de aves, y se designaron puntos de observación de aves (n = 24) para cada par de parcelas entre control y exclusión (Figura 1a). Se realizaron dos censos de aves durante el período principal de reproducción (i.e. con mayor detectabilidad): el primero entre el 2–4 de mayo y el segundo entre el 15–17 de mayo. Además, se clasificaron las especies de aves como insectívoras funcionales, es decir, aquellas que son insectívoras durante el período reproductivo y que forrajean principalmente en hojas o cazan en vuelo.
Para los muestreos de murciélagos, se realizaron registros acústicos entre el 17–19 de mayo y el 12–14 de julio, durante 5 h en tres noches consecutivas, comenzando 30 min antes de la puesta del sol. Se instalaron dispositivos acústicos de espectro completo AudioMoth (n = 24) para registrar los ultrasonidos de murciélagos (Hill et al., 2018) en los mismos puntos usados para la observación de aves (Figura 1a).
La temperatura promedio de los sitios de muestreo fue de 15,9 ± 0,2 °C en mayo y 27,2 ± 0,2 °C en julio. Para las grabaciones de llamadas, la frecuencia de muestreo fue de 256 kHz; la ganancia se configuró en nivel medio; las grabaciones duraban 295 s con 5 s de pausa para guardar los archivos. Se buscaron automáticamente las llamadas de murciélagos con el programa Bat Detective y se recortaron las secuencias con un script propio en el entorno estadístico R 4.4.2 (R Core Team, 2024). Se definió una secuencia como una serie de llamadas de ecolocalización con intervalos interpulso menores a 3 s. Finalmente, validamos manualmente las identificaciones automáticas a nivel de especie en el software Kaleidoscope Pro (Wildlife Acoustics, Inc.) y se definió la actividad de murciélagos como el número de intervalos de 5 s que contenían llamadas.
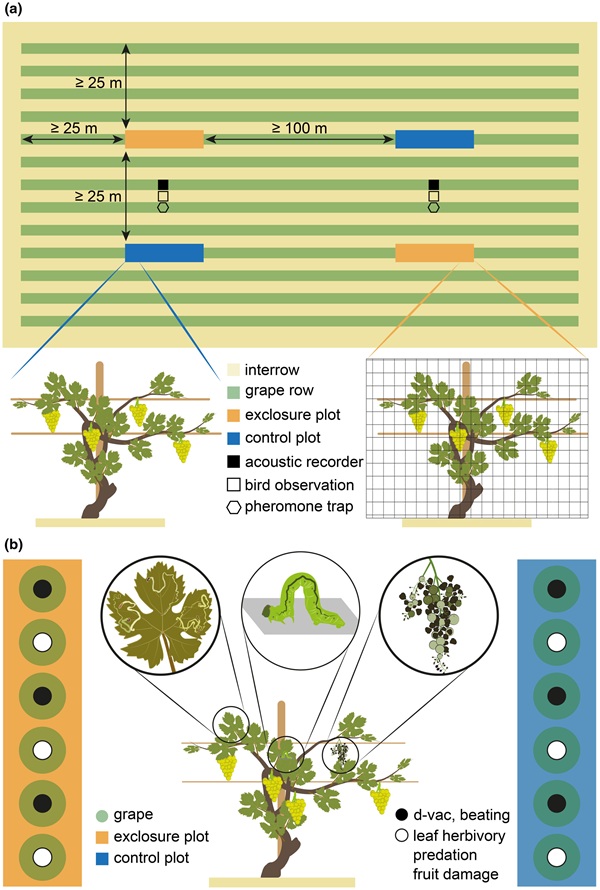
Para recolectar individuos de Lobesia botrana, se utilizaron trampas con feromonas específicas (CSalomon® RAG, Plant Protection Institute, Hungría). Se instalaron las trampas a nivel del dosel, cerca de los puntos de observación de aves y murciélagos (n = 24, Figura 1a), y se recolectaron polillas durante 7 días en tres ocasiones: 15–21 de mayo, 10–16 de julio y 21–27 de agosto. En los mismos periodos recolectamos artrópodos del dosel mediante los métodos de golpeteo y aspiración D-Vac (Basset et al., 1997) en las plantas impares (1, 3, 5) de cada parcela (Figura 1b). El material obtenido por ambos métodos y de las tres plantas de cada parcela se procesó en conjunto (n = 48). Se contaron e identificaron los individuos hasta el nivel taxonómico más bajo posible, cuando era factible clasificarlos en grupos funcionales (herbívoros y depredadores).
Se midió herbivoría foliar, daño en frutos y depredación en las plantas pares (2, 4, 6) de cada parcela (Figura 1b). Se evaluó el daño en las bayas por artrópodos -principalmente perforaciones de larvas de L. botrana– calculando la proporción de bayas dañadas en cinco racimos seleccionados al azar por planta (n = 48 × 3 × 5 = 720), antes de la cosecha y del retiro de las jaulas a finales de agosto. Simultáneamente, se inspeccionó visualmente la herbivoría foliar en diez hojas escogidas al azar por planta (n = 48 × 3 × 10 = 1440) y se calculó la proporción de superficie dañada por insectos herbívoros. Se investigó la depredación por artrópodos mediante un experimento con presas centinela a mediados de mayo: se fijaron orugas artificiales en cinco hojas seleccionadas al azar por planta (n = 48 × 3 × 5 = 720) y se expusieron durante 7 días para luego inspeccionar las marcas de depredación en el laboratorio. El trabajo de campo se llevó a cabo con el consentimiento de los propietarios de los viñedos y no requirió permisos específicos ni aprobaciones éticas.
Análisis de datos
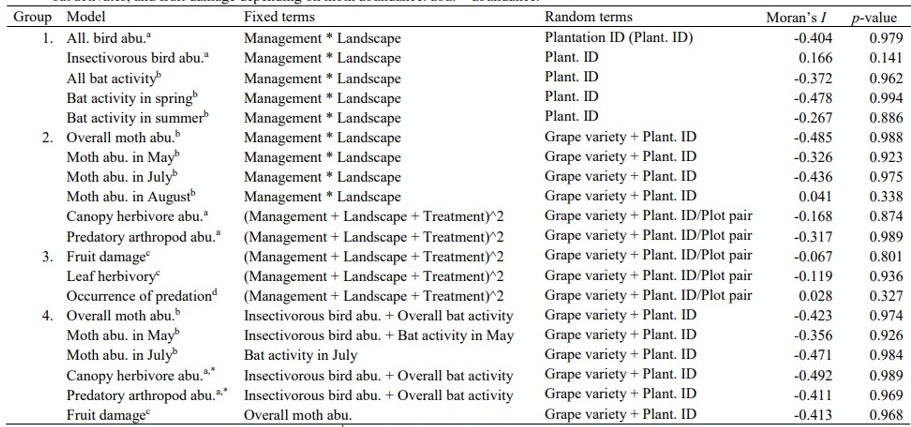
Los investigadores crearon cuatro grupos de modelos lineales generalizados de efectos mixtos para probar el efecto de:
- El manejo local del viñedo (orgánico vs. IPM), el paisaje (forestado vs. no forestado) y su interacción sobre la abundancia total e insectívora de aves, así como sobre la actividad total y estacional (primavera y verano) de murciélagos.
- Manejo, proximidad al bosque y tratamiento (control vs. exclusión; excepto polillas) y sus interacciones sobre la abundancia total y temporal de polillas y de artrópodos del dosel.
- Manejo, proximidad al bosque, tratamiento y sus interacciones sobre la herbivoría foliar, daño en frutos y ocurrencia de depredación.
- Relaciones funcionales, es decir: (a) el efecto de la abundancia de aves y actividad de murciélagos sobre la abundancia total de artrópodos del dosel y de polillas (total y temporal en mayo y julio), y (b) el efecto de la abundancia de polillas sobre el daño en frutos.
Para la estructura de los modelos completos y términos aleatorios incluidos, véase la Tabla S3.
Todas las analíticas se realizaron en el entorno R 4.4.2. Los modelos se ajustaron usando las funciones glmer (regresión de Poisson: abundancia de aves y artrópodos del dosel; regresión binomial: ocurrencia de depredación) y glmer.nb (regresión binomial negativa: actividad de murciélagos y abundancia de polillas) del paquete lme4 (Bates et al., 2015), y glmmTMB (regresión beta ordenada: herbivoría foliar y daño en frutos) del paquete glmmTMB (Brooks et al., 2017). Realizamos una selección automática de modelos basada en AICc con el paquete MuMIn (Bartoń, 2024) y aplicamos promedio de modelos cuando ΔAICc < 2. Las pruebas de los modelos se realizaron con test χ² de Wald tipo II mediante la función Anova del paquete car (Fox & Weisberg, 2019).
Cálculo del rendimiento
Para determinar el papel de aves y murciélagos en la influencia sobre el rendimiento del cultivo, se siguió la metodología de Rodríguez-San Pedro et al. (2020). Primero, se calculó la diferencia entre los porcentajes promedio generales de bayas dañadas observados en parcelas control y de exclusión (para los mismos racimos usados en el análisis de daño en frutos). Luego, se calculó el rendimiento promedio (kg/ha) considerando los datos de los viñedos estudiados, obtenidos directamente de los propietarios. Finalmente, se multiplicó este valor promedio de rendimiento por la diferencia en el daño promedio de bayas entre los tratamientos.
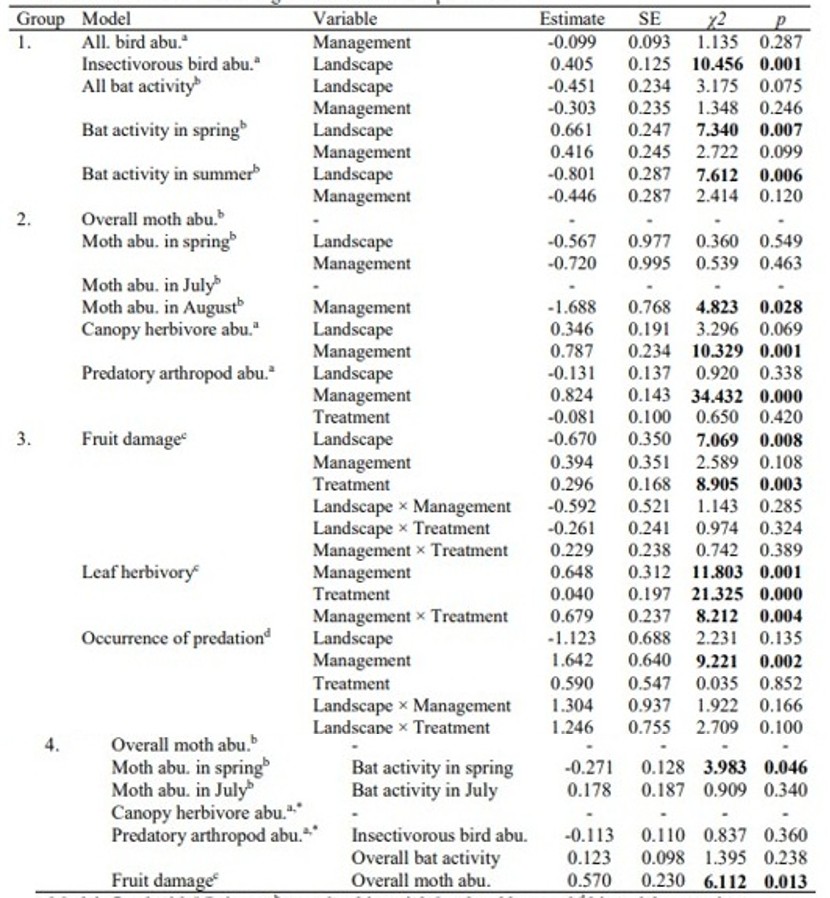
Modelo: ajustado con Poisson (a), binomial negativa (b), beta ordenada (c) y regresión binomial (d). Estimación: valores positivos para paisaje, manejo y tratamiento indican números más altos en viñedos en paisajes forestales, aquellos con manejo orgánico y parcelas de exclusión, respectivamente.
Solo se consideraron los datos de las parcelas de control.
Resultados
Se observaron 1.068 aves pertenecientes a 50 especies, de las cuales 265 individuos de 17 especies eran insectívoros funcionales (Tabla S5), y se registraron 3.412 pases de murciélagos que correspondieron a nueve taxones identificados (Tabla S6). El análisis de la abundancia total de aves y la actividad de murciélagos no mostró efectos significativos de ninguna de las variables.
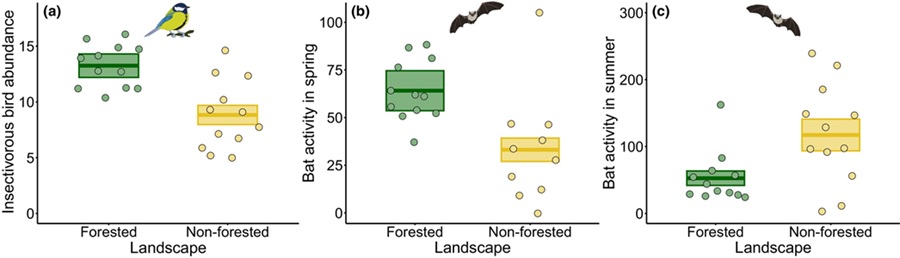
Sin embargo, se encontró un efecto significativo del paisaje sobre las aves insectívoras, con mayor número en viñedos situados en paisajes forestados en comparación con los ubicados en paisajes no forestados (+50%, χ² = 10,456, p = 0,001; Figura 2a). Además, la actividad de murciélagos en primavera fue mayor en viñedos forestados que en los no forestados (+127%, χ² = 7,340, p = 0,007; Figura 2b), mientras que su actividad en verano fue mayor en viñedos situados en paisajes no forestados (+124%, χ² = 7,612, p = 0,006; Figura 2c). El manejo del viñedo no tuvo un efecto significativo sobre las densidades de aves ni la actividad de murciélagos (Tabla S4).
Recolectamos 3.601 individuos de Lobesia botrana, 256 artrópodos herbívoros del dosel y 396 artrópodos depredadores (Tablas S7 y S8). No encontramos efectos significativos del manejo ni del paisaje sobre la abundancia de L. botrana, excepto en agosto, cuando fue significativamente mayor en viñedos IPM que en orgánicos (+165,9%, χ² = 4,823, p = 0,028; Figura 3a).
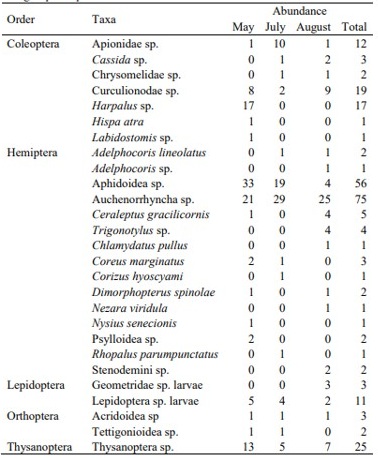
El manejo del viñedo afectó significativamente la abundancia de artrópodos del dosel, con más artrópodos herbívoros y depredadores en los viñedos orgánicos que en los IPM (herbívoros: +124,1%, χ² = 10,329, p = 0,001; depredadores: +130%, χ² = 34,432, p = 0,000; Figura 3b,c). Las densidades de artrópodos del dosel no se vieron afectadas por la proximidad al bosque ni por el tratamiento (Tabla S4).
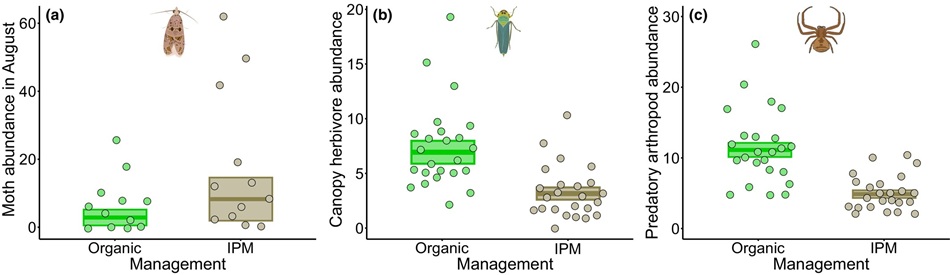
El daño promedio en los frutos fue significativamente mayor en viñedos situados en paisajes no forestados que en los forestados (diferencia de 2,1%, χ² = 7,069, p = 0,008; Figura 4a) y también en las parcelas de exclusión, donde los enemigos naturales no tenían acceso a las plantas, en comparación con las parcelas control (χ² = 8,905, p = 0,003; Figura 4b). En las parcelas de exclusión, el 3,4% de las bayas estaban dañadas, frente al 2,2% registrado en las parcelas control, lo que supone una diferencia de 1,2%, es decir, la proporción de bayas dañadas aumentó un 54,5% con la exclusión.
En los viñedos estudiados, el rendimiento promedio fue de 6.085,4 kg/ha (IPM: 8.750; orgánico: 3.420,8). Considerando estos valores, estimamos que la exclusión de aves y murciélagos redujo el rendimiento en 73,1 kg/ha en promedio (IPM: 105; orgánico: 41,1).
Se encontró un efecto significativo de interacción entre manejo y tratamiento sobre la herbivoría foliar, siendo significativamente mayor en las uvas excluidas que en las control en los viñedos orgánicos (diferencia de 1,3%, χ² = 8,212, p = 0,004; Figura 4c). Además, el manejo orgánico afectó significativamente y de manera positiva la ocurrencia de depredación mediada por artrópodos sobre las uvas (+104,5%, χ² = 9,221, p = 0,002; Figura 4d; Tabla S4).
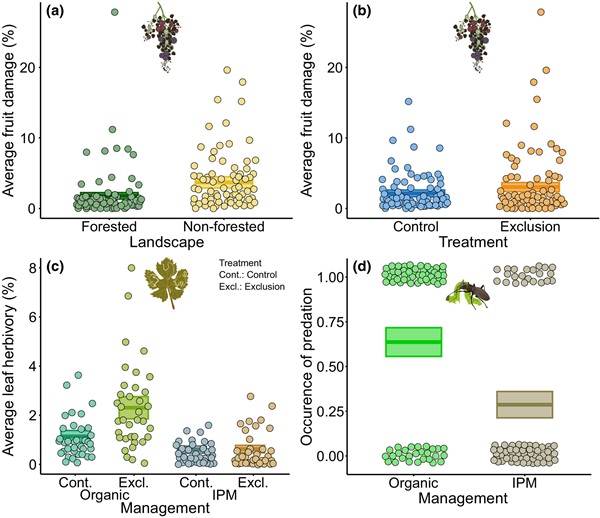
Con respecto a las relaciones funcionales, la abundancia de polillas disminuyó significativamente con el aumento de la actividad de murciélagos en primavera (36,7% menos de abundancia al incrementarse la actividad de 0 a 100; χ² = 3,983, p = 0,046; Figura 5a). Además, el daño promedio en los frutos aumentó significativamente con el incremento de la abundancia total de polillas (66,7% más de daño al aumentar la abundancia de 0 a 100; χ² = 6,112, p = 0,013; Figura 5b). Para más efectos no significativos retenidos tras la selección de modelos, véase Tabla S4.
Discusión
En este estudio experimental de exclusión, se investigó el papel de aves y murciélagos en la regulación de las abundancias de artrópodos y las funciones ecosistémicas asociadas en viñedos con manejos locales contrastantes (orgánico vs. IPM) y heterogeneidad paisajística (forestado vs. no forestado). Además de la escasez de experimentos de exclusión en agrosistemas permanentes europeos, este estudio es el primero en investigar el potencial de control biológico de estos depredadores vertebrados en viñedos considerando simultáneamente efectos de manejo y paisaje. Se encontró que el manejo del viñedo sólo afectó a los artrópodos del dosel, alcanzando mayores abundancias y contribuyendo a una mayor herbivoría foliar y presión de depredación en viñedos orgánicos que en IPM. Al mismo tiempo, los paisajes forestados tuvieron un efecto positivo sobre aves insectívoras y murciélagos durante el periodo reproductivo. También se demostró que estos depredadores vertebrados actúan como agentes de control de plagas, pudiendo reducir la herbivoría y aumentar los beneficios económicos.
Hipótesis 1: Efecto positivo de los paisajes forestados sobre aves y murciélagos
Se encontraron efectos significativos de los paisajes forestados sobre la abundancia de aves y la actividad de murciélagos, aunque este último efecto dependió de la estación. Coincidiendo con estudios previos, las aves insectívoras se benefician de la proximidad y disponibilidad de bosques, que proporcionan sitios de anidación protegidos, estructuras de forrajeo y abundantes fuentes de alimento. Los bosques caducifolios de esta región húngara, dominados por el roble común (Quercus petraea), favorecen comunidades de artrópodos abundantes y diversas, como fuente de alimento para aves insectívoras que recolectan insectos de las hojas.
Los paisajes forestados pueden incrementar significativamente la actividad de forrajeo de los murciélagos. Estos hábitats son especialmente importantes en primavera, ya que ofrecen diversidad de presas y microhábitats de refugio para muchas especies europeas. La preferencia por sitios menos forestados en verano puede deberse a que los murciélagos ya no están restringidos a áreas boscosas; para entonces, los juveniles han abandonado los sitios de maternidad, lo que les permite forrajear durante períodos y distancias más largas. Aunque no se exploró en este estudio, la proporción relativamente alta de áreas agrícolas abiertas en paisajes no forestados también podría ofrecer zonas de caza ricas en alimento para algunas especies que buscan presas en espacios abiertos o bordes de cultivo en esta estación.
Contrario a esta hipótesis, el manejo no afectó directamente a aves ni murciélagos, presentando densidades y actividades similares en viñedos orgánicos e IPM. Este patrón puede atribuirse a que aves y murciélagos tienen la mayor capacidad de dispersión entre los grupos taxonómicos estudiados, de modo que las características del paisaje podrían ser más determinantes que los factores locales.
Hipótesis 2: Murciélagos como reguladores de poblaciones de polillas
Se encontró que un aumento en la actividad de murciélagos afectó negativamente la abundancia de L. botrana en primavera, pero no impactó las densidades de artrópodos del dosel. Los murciélagos son depredadores voraces y oportunistas, con respuestas numéricas ante el incremento de presas, incluyendo lepidópteros. En particular, L. botrana constituye una parte importante de la dieta de murciélagos en regiones vitivinícolas europeas, influyendo en su actividad de caza. Esta especie fue la más abundante entre los artrópodos estudiados y presentó la mayor densidad en primavera (1.791 de 3.601 individuos capturados), por lo que pudo ser fuente primaria de alimento para los murciélagos en esta época.
En contraste, los artrópodos del dosel presentaron densidades relativamente bajas y no representaron un recurso de presas suficientemente concentrado para los murciélagos, siendo afectados principalmente por el manejo, con mayores abundancias en viñedos orgánicos debido a la liberación de insecticidas (efecto directo) y herbicidas (efecto indirecto por menor diversidad y complejidad estructural de la vegetación herbácea).
Aunque se esperaría mayor abundancia de L. botrana en viñedos alejados de bosques, la falta de este patrón podría explicarse por la capacidad de los bosques densos y sombreados de mitigar periodos cálidos y secos críticos para el desarrollo de L. botrana, especialmente en primavera. Esto, junto con la mayor actividad de aves y murciélagos en paisajes forestados y posiblemente mayor exposición a pesticidas en paisajes agrícolas simplificados, podría generar patrones de densidad de polillas similares entre viñedos con distintas características paisajísticas.
Hipótesis 3: Servicios ecosistémicos proporcionados por enemigos naturales
Se observó mayor daño foliar y en frutos en uvas excluidas, lo que indica el papel potencial de aves y murciélagos en la reducción de la herbivoría y el aumento del rendimiento del cultivo. Asimismo, encontramos menor daño en frutos en viñedos situados en paisajes forestados, preferidos por estos depredadores durante el período reproductivo. Los resultados coinciden con estudios previos que muestran que los murciélagos proporcionan servicios ecosistémicos esenciales, como la reducción de daños a las plantas y el aumento del beneficio económico para los agricultores. Además, los murciélagos regulan plagas dañinas para los frutos no solo en bosques, sino también en áreas agrícolas adyacentes.
Un metaanálisis global mostró que las aves también son reguladores importantes de poblaciones de plagas, reduciendo significativamente el daño en plantaciones leñosas, incluidos los viñedos. Sin embargo, no se detectó una relación negativa entre la abundancia de aves y L. botrana, lo que sugiere que las aves podrían usar principalmente los bosques caducifolios circundantes y sus bordes como hábitats de forrajeo, impidiendo potencialmente la dispersión de plagas hacia los viñedos. Se requerirían muestreos de artrópodos más frecuentes (especialmente en primavera) y observaciones de aves posteriores a la temporada reproductiva, incluyendo bordes de plantación, para establecer relaciones más sólidas entre la actividad de aves y la densidad de plagas.
No se encontró evidencia de que aves y murciélagos disminuyan la presión de depredación ejercida por artrópodos. La presión de depredación mediada por artrópodos fue mayor en viñedos orgánicos que en IPM y no se vio afectada por la exclusión. Esto puede explicar la baja abundancia de L. botrana en viñedos orgánicos en agosto, ya que las arañas, los artrópodos del dosel más abundantes en nuestro estudio, son depredadores clave de estas polillas.
Perspectivas aplicadas
Este estudio destaca que asegurar la proximidad (30 m en promedio) y la proporción adecuada (25%–40% según la escala espacial) de parches de bosque caducifolio alrededor de los viñedos promueve aves y murciélagos, así como la regulación de plagas y el aumento del rendimiento. Estos parches son hábitats importantes durante el período reproductivo, especialmente si contienen una alta proporción de especies arbóreas nativas (p. ej., Quercus petraea), con comunidades de artrópodos abundantes y diversas, garantizando recursos de forrajeo valiosos.
El valor de conservación de estos hábitats puede incrementarse mediante prácticas de manejo menos intensivas, fomentando vegetación heterogénea y árboles grandes para proporcionar microhábitats adecuados para anidación y refugio. Considerando las características del paisaje de los viñedos estudiados, también es importante la conectividad de elementos leñosos, ya que redes de parches y elementos lineales (grupos de árboles, matorrales y setos) pueden ofrecer hábitats óptimos de forrajeo y “escalones” para depredadores vertebrados incluso en paisajes simplificados y menos forestados.
Estos resultados sugieren que el control biológico mediado por aves y murciélagos puede potenciarse facilitando la colonización de artrópodos beneficiosos en los viñedos. Esto subraya la necesidad de aumentar el número de viñedos.
Autores: Dávid Korányi, András Báldi y Péter Batáry (Institute of Ecology and Botany Hungría); Sándor Zsebők (Eötvös Loránd University, Budapest, Hungría), Mattia Brambilla (University of Milan, Italia); y Máté Varga (Varga Winery, Badacsonyörs, Hungría).
Fuente: British Ecological Society
