

El cultivo de la vid en altitudes elevadas ha ido adquiriendo importancia en las últimas décadas como opción de calidad para producir vinos en regiones cálidas por el avance del cambio climático. Numerosos países productores europeos y sudamericanos están experimentando con el cultivo de la uva de altura desde una perspectiva innovadora.
Los investigadores argentinos Leonardo Arias, Federico Berli, Ariel Fontana y Patricia Piccoli (del Grupo de Bioquímica Vegetal, Instituto de Biología Agrícola de Mendoza, Conicet, Mendoza) y Rubén Bottini (del Instituto Argentino de Veterinaria, Ambiente y Salud de la Universidad Juan Agustín Maza, Mendoza), publicaron recientemente en la revista científica Frontiers in Plant Science este completo relevamiento de los estudios, ensayos en el campo y literatura científica sobre este tema, el que a continuación replicamos.
Ciertamente, la calidad de la uva depende de aspectos que involucran el medio ambiente y los enólogos necesitan comprender mejor cómo los cambios que ya están aquí, producidos per el calentamiento global, afectarán al crecimiento de las bayas.
El incremento en la temperatura del aire, la concentración de CO2 y la escasez de agua son escenarios sobre los que la comunidad científica está de acuerdo en que incluso reduciendo enormemente las emisiones, las temperaturas se mantendrán muy elevadas hasta después del año 2100.
En muchas áreas de actividad y sociales no se termina de asimilar, pero en el caso de la enología, ya se están implementando planes para desplazar los cultivos a estas zonas más elevadas. Pero, ¿cuánto hay que subir en la ladera? ¿Cuál es la altitud mínima que afecta de forma diferencial al crecimiento y el desarrollo de la vid?
Este artículo, con pruebas de campo realizadas en Argentina, Suiza e Italia, resume los efectos de las variables climáticas globales relacionadas con la gran altitud en la fisiología de la vid y la composición del vino, para una mejor evaluación del posible establecimiento de viñedos a gran altitud en escenarios de cambio climático.
Es interesante que a elevadas altitudes, las uvas muestren mayor contenido de antocianinas y mayor acidez, contrarrestando los efectos negativos de una maduración acelerada. Ésta y otras ventajas son objeto de estudio en esta revisión.
La altitud del viñedo y su relación con el vino obtenido
El cultivo de la vid para la vinificación depende en gran medida del clima. Normalmente, requiere temperaturas medias del aire que oscilan entre 12°C y 22°C durante el ciclo de crecimiento, al menos 700–900 μmol fotones m−2 s−1 de radiación solar, alrededor de 600–800 mm de lluvia o riego suficiente y una helada -temporada de crecimiento libre (Palliotti y Poni, 2016; Zapata et al., 2016; Ateş y Uysal, 2017). Esto generalmente restringe el cultivo de la uva más allá de los 50° de latitud N y los 40° de latitud S (Kenny y Shao, 1992; Cabré et al., 2016).
La viticultura de altura ha ido adquiriendo gran importancia en las últimas décadas debido a su potencial para producir vinos de alta calidad en regiones más cálidas, ya sea por la naturaleza de su geografía o por un avance del cambio climático. Ciertos países, como Argentina, Brasil, China, Italia, Portugal y Turquía, han participado en una expansión tanto del cultivo de la uva de altura como de su estudio científico.
La Tabla 1 (ver más abajo) resume la literatura actual sobre la altitud del viñedo y su efecto en varias características de la vid y el vino. En particular, estos países cultivan a altitudes superiores a la media de las regiones vitivinícolas (ca. 300 m snm; Gladstones, 2011). Lo que se considera viñedo de altura en la literatura actual va desde los 350 m s.n.m. en el Valle del Duero (Oliveira et al., 2004) a 2.900 m s.n.m. en el suroeste de China (Xing et al., 2016). Esto nos permite definir la altitud elevada en términos relativos, señalando el cambio mínimo de altitud que afecta diferencialmente al crecimiento y desarrollo de la vid, así como a la calidad del vino, para una región determinada.
Ver Tabla 1: AQUÍ
Según Körner (2007), las variables ambientales que cambian con la altitud se pueden agrupar en dos categorías: las derivadas de la altitud per se, como la temperatura, la presión atmosférica y la turbidez del cielo; y las relacionados con la proximidad a terrenos montañosos, como la humedad, el viento y la geología. Esta revisión se enfoca en las primeras variables, en virtud de ser globalmente comparables, mientras que las segundas son demasiado específicas para un sitio como para resumirlo.
Cambio climático: ¿Qué está cambiando exactamente?
Según el sexto informe de evaluación del Panel Intergubernamental sobre el Cambio Climático (IPCC, por sus siglas en inglés), las últimas cuatro décadas han sido secuencialmente más cálidas que cualquier otra década desde 1850 (IPCC, 2021).
En particular, la temperatura de la superficie terrestre en las dos primeras décadas del siglo XXI fue 1,59 °C más alta que entre 1850 y 1900. Los extremos cálidos se han vuelto más frecuentes e intensos, mientras que los extremos fríos se han vuelto menos frecuentes y severos desde 1950. Además, desde 1750, las concentraciones de CO2 han aumentado continuamente debido a las actividades humanas, alcanzando promedios anuales de 410 ppm. De manera alarmante, es muy probable que la mayoría de estos aspectos persistan durante muchos siglos, incluso cuando se detuvieran las emisiones de CO2 (Gillett et al., 2011; IPCC, 2013).
El IPCC ha realizado varias proyecciones de cambio climático a partir de un conjunto de escenarios de impacto antrópico, desde los más optimistas hasta los más pesimistas, que darían como resultado diferentes magnitudes de emisiones, o Vías de Concentración Representativas (RCPs). Estos incluyen un aumento en las temperaturas superficiales medias globales de 1,7 °C (RCP2,6) a 4,8 °C (RCP8,5) para el período 2081-2100 en relación con 1986-2005, un aumento en la concentración de CO2 atmosférico que alcanza los 421 ppm (RCP2 .6) a 936 ppm (RCP8.5) para el año 2100, y una certeza virtual de temperaturas extremas calientes más frecuentes y menos frías (IPCC, 2013).
Dado este escenario, se esperan impactos importantes en los sistemas rurales, incluidos cambios en las áreas de producción de cultivos alimentarios, así como estrategias de adaptación, como prácticas culturales y variedades de cultivos más adecuadas (IPCC, 2014; Fraga, 2020).
Cambio climático y fenología de la vid
Webb et al. (2012) llevaron a cabo un estudio de factores climáticos de temperatura y factores no relacionados con la temperatura, acerca de la madurez de la uva para vino en el sur de Australia. La fecha de cosecha estuvo consistentemente mejor representada por las temperaturas promedio durante la temporada de crecimiento en todos los sitios, seguida por una menor humedad del suelo. Los turnos de entre 4 y 11 días se atribuyeron al calentamiento. La reducción de la humedad del suelo adelantó la madurez independientemente del aumento de la temperatura, aunque las recientes tendencias de sequía en Australia se han atribuido al cambio climático antropogénico.
Arrizabalaga-Arriazu et al. (2020b) determinaron la respuesta fenológica de los clones de tempranillo a la temperatura del aire y la concentración de CO2 prevista para 2100. En general, entre los clones, la temperatura elevada redujo el tiempo total necesario para alcanzar la madurez de la fruta. El CO2 no tuvo efectos significativos en la fenología vegetal, con excepción del clon RJ43, que fue el más afectado por los escenarios de cambio climático. Esto reveló la importancia del material vegetal y la variabilidad genética para la adaptación climática.
Ruml et al. (2016) examinaron las fechas históricas de inicio de brotación, floración, pinta y cosecha de 20 cultivares de uva para vino durante el período 1981–2007. Aunque todos los estados fenológicos avanzaron significativamente, el mayor efecto térmico se observó desde el inicio de la floración hasta el inicio del pinta, mientras que el menor se presentó durante el período de maduración. Las tendencias fueron − 0,4 día/año para floración, −0,7 días/año para inicio de pinta y −0,6 día/año para fecha de cosecha. Los autores también encontraron efectos no lineales del calentamiento, que explican hasta el 26 % de la variación en el momento fenológico, atribuido a un cambio del período de maduración hacia condiciones más cálidas a principios de la temporada debido a la floración y el envero precoces.
En un estudio con la variedad tempranillo en el sur de Australia, sorprendentemente, un aumento de 1 °C en la temperatura media durante el inicio de la brotación, la floración o el envero fue suficiente para adelantar el tiempo medio de cosecha en 7,4 días.
En entornos experimentales de invernadero que simulan condiciones de temperatura, CO2 y déficit hídrico previstos para finales de siglo, (Martínez-Lüscher et al. 2016a,b) se advirtió un avance significativo en la brotación, la floración y la madurez de la baya en tempranillo tinto y blanco. Las condiciones con 700 ppm CO2 y 28°C dieron como resultado un inicio más temprano del envero de aproximadamente 9 días y un tiempo de cosecha (22 °brix) de alrededor de 23 días. En general, el efecto de la temperatura en la madurez estuvo entre −0,90 días/°C y − 2,03 días/°C en todos los años y variedades, mientras que el efecto del CO2 para todos los años y variedades fue un avance de los tiempos fenológicos en 6,75 días para la temperatura ambiente y 4.06 días para T + 4°C.
Hall et al. (2016) modelaron eventos de brotación y cosecha con datos de temperatura interpolados de 1975 a 2004, y proyectaron condiciones más cálidas en diferentes escenarios de emisiones en las regiones vitivinícolas de Australia. Este estudio sugiere que la brotación y la madurez de la fruta ocurrirán antes en todas las regiones australianas, lo que aumentará la acumulación de calor poscosecha, especialmente en las regiones más frías. Esto provocaría una acumulación de carbohidratos y reservas de nutrientes en las estructuras perennes, lo que probablemente mejoraría el vigor y el crecimiento vegetativo en el año siguiente.
Ramos (2017) realizó predicciones fenológicas de tres variedades de uva chardonnay, macabeo y parellada para un clima mediterráneo (España) sin riego para los años 2030, 2050 y 2070 según RCP4.5 y RCP8.5. El estudio sugirió un inicio más temprano de todas las etapas fenológicas, principalmente envero y cosecha, con un acortamiento tanto del tiempo fenológico como de los intervalos entre fases, lo que puede afectar la calidad de la uva. Esto también fue cierto para la variedad tempranillo (Ramos y de Toda, 2020). Además, una reducción del agua disponible debido a una mayor evapotranspiración puede acentuar el avance de algunas etapas fenológicas, como el envero. El envero y la cosecha podrían avanzar hasta 12 y 20 días para 2070 bajo el escenario RCP4.5, y 23 y 28 días bajo el escenario RCP8.5.
Cambio climático y fisiología de la vid
Schultz y Hofmann (2016) revisaron cómo el aumento de CO2 a alrededor de 560 ppm causará una disminución general de la humedad relativa del aire y una mayor demanda de evaporación por medio de una reducción de la conductancia estomática de alrededor del 20 %. Con el doble de CO2, la conductancia estomática se reduciría en alrededor de un 40 %, lo que a su vez reduciría la transpiración en un 20 %, la evapotranspiración entre un 10 % y un 14 %, y un menor enfriamiento por evaporación debería resultar en una temperatura más alta experimentada por las superficies de las plantas.
En zonas sin riego, tales niveles de déficit no serían deseables en términos de calidad del vino. Además del efecto indirecto del CO2 en la temperatura global, el CO2 en sí mismo debería ser beneficioso para el crecimiento de la vid, aumentando la producción de biomasa con una menor pérdida de agua y, en última instancia, aumentando la concentración de azúcar en la fruta y disminuyendo los niveles de acidez.
Salazar-Parra et al. (2012) evaluaron la influencia de las condiciones de CC (temperatura elevada y CO2, y déficit hídrico moderado) sobre la condición antioxidante de las hojas de tempranillo desde el envero hasta la madurez. La temperatura ambiente elevada (28°C) y el CO2 (700 ppm) presentaron menor peroxidación de lípidos de membrana y concentraciones de ROS (como H2O2) en las hojas, particularmente en condiciones de buen riego en comparación con el déficit hídrico. Se atribuyó a aumentos en la fijación de carbono de la fotosíntesis. Las hojas bajo estrés hídrico moderado y las condiciones ambientales presentes (375 ppm y 24°C) fueron más susceptibles al daño oxidativo, lo que indica que el sistema antioxidante fue menos capaz de aliviar el daño (Salazar-Parra et al., 2012).
Arrizabalaga-Arriazu et al. (2020b) estudiaron la respuesta fisiológica de cinco clones de tempranillo a la temperatura y CO2 previstos para el año 2100 en un experimento de invernadero. La fotosíntesis neta se estimuló inicialmente en el envero, pero se produjo una regulación a la baja de la fotosíntesis debido al CO2 elevado combinado con la alta temperatura que causa la limitación de nitrógeno. Se sabe ampliamente que la concentración de CO2 se correlaciona negativamente con la conductancia de los estomas (Lammertsma et al., 2011).
Cambio climático y bioquímica de la uva
Martínez-Lüscher et al. (2016b) encontraron que el CO2 elevado y el aumento de la temperatura redujeron la proporción de antocianinas a azúcar en las bayas de uva, junto con un inicio más temprano del envero y sólidos solubles totales más altos.
Van Leeuwen y Destrac-Irvine (2017) se centraron en el efecto de la temperatura, los rayos UV-B y el estado del agua en la viticultura. El efecto combinado del aumento de las temperaturas y la fenología avanzada provocó más azúcar y menos ácidos orgánicos en las bayas, y alteró la composición de los metabolitos secundarios, en particular los precursores del aroma.
En un estudio de campo destinado a evaluar el efecto de +20% de CO2 suplementado en vinos riesling y cabernet sauvignon, Wohlfahrt et al. (2021) reportaron un efecto nulo o pequeño del CO2 en los ácidos orgánicos y azúcares en el mosto de uva, pero en la proporción de ácido tartárico a ácido málico, el ácido tartárico se vio favorecido por ambos cultivares. Hubo, sin embargo, una menor concentración de antocianinas debido al efecto indirecto del aumento del tamaño de la baya del CO2.
Los análisis de componentes principales revelaron que el efecto de la cosecha fue en general más fuerte que el tratamiento con CO2 elevado en +20 % (Wohlfahrt et al., 2021). Del mismo modo, Arrizabalaga-Arriazu et al. (2020a) estudiaron la composición del mosto y los antocianos totales de la piel para clones de tempranillo en las condiciones climáticas previstas para 2.100.
Se prevé que los aumentos en el CO2 y la temperatura promedio del aire reduzcan el tiempo de maduración y mejoren la acumulación de azúcar y la descomposición del ácido málico, al tiempo que retrasan la madurez de los aminoácidos.
Aunque no se prevé que la concentración de antocianinas cambie en estas condiciones, podría haber una reducción de la proporción entre antocianinas y azúcares. Se cree que el grado en que la temperatura alta afecta la proporción de antocianina a azúcar depende del viñedo, debido a la diferente sensibilidad de la antocianina de la baya a los rangos críticos de temperatura (Fernandes de Oliveira et al., 2015).
Además, Ramos y de Toda (2020) proyectaron escenarios de cambio climático tanto optimistas como pesimistas para la composición de la uva, y estimaron valores decrecientes de acidez total en tres zonas analizadas a partir de los cambios de temperatura proyectados.
Cambios en la distribución vitivinícola
La magnitud en la que los factores ambientales restringen la viticultura para la producción de uva difiere considerablemente de una región a otra (Schultz y Hofmann, 2016). Asimismo, los impactos del cambio climático variarán en consecuencia. Se espera que los sitios donde la viticultura se caracteriza por temporadas de crecimiento cortas o temperaturas de verano bajas, como aquellos ubicados en latitudes o elevaciones más altas, se vuelvan progresivamente más cálidos bajo las condiciones climáticas futuras, lo que permitirá que se cultive de manera confiable una selección más amplia de cultivares (Mosedale et al. , 2016).
Las regiones productoras de uva premium actuales pueden sufrir reducciones importantes en la calidad debido al aumento proyectado de la temperatura y la menor disponibilidad de agua.
El índice de Huglin (HI) ha demostrado ser una herramienta eficaz para la zonificación vitivinícola y, por lo tanto, se ha aplicado ampliamente (Jones et al., 2010). Fraga et al. (2013) analizaron los patrones de HI simulados para el período 2041-2070 en Europa y mostraron un desplazamiento hacia el norte de áreas de alta idoneidad para la viticultura. Las regiones del sur de Europa, por otro lado, tendrán una idoneidad reducida principalmente debido al aumento de la sequía, y grandes áreas de Europa central y occidental se volverán más idóneas debido a las condiciones térmicas favorables (Fraga et al., 2013).
Mosedale et al. (2015) sugirieron que, en escenarios climáticos futuros, incluso con bajas emisiones, habrá una gama mucho más amplia de cultivares adecuados para cosechas de uvas de vinificación de alta calidad en regiones de clima frío. El sureste de Inglaterra, por ejemplo, actualmente es adecuado para producir vino espumoso de alta calidad, que requiere uvas particularmente ácidas, pero en escenarios futuros podría permitir cosechas confiables de chardonnay o pinot noir (Mosedale et al., 2015).
Lorenzo et al. (2016) evaluaron cuatro modelos climáticos regionales (MCR) diferentes para España, utilizando tres índices bioclimáticos para la viticultura (Huglin y Branas, Winkler y Bernon y Levadoux). Todas las MCR coinciden en una mayor acumulación de calor en el centro y sur de España, lo que repercutiría negativamente en la calidad del vino (Lorenzo et al., 2016). Además, se prevé que, a finales del siglo XXI, el sur de España ya no sea adecuado para la producción de vino de alta calidad debido al obstáculo térmico para el crecimiento de la vid (Lorenzo et al., 2016). Por otro lado, el noroeste de España podría beneficiarse del calentamiento proyectado, aumentando la calidad potencial del vino (Lorenzo et al., 2016). Se observa que los diferentes índices bioclimáticos no están de acuerdo sobre el tamaño del cambio futuro proyectado.
Entre 2075 y 2099, en Cuyo habrá desplazamientos al sur y oeste
Cabré et al. (2016) también analizaron la simulación de un modelo climático regional a través de varios índices bioclimáticos, para la región de Cuyo en Argentina, donde se produce la mayor parte de los vinos del país. Las proyecciones predicen un desplazamiento de áreas adecuadas hacia el sur, a latitudes más altas, y hacia el oeste, a latitudes más altas, para acomodarse para el cambio de temperatura a finales de siglo (2075-2099; Cabré et al., 2016; Cabré y Nuñez, 2020). Así, bajo los escenarios proyectados, la principal región vitivinícola de la Argentina actual enfrentaría grandes desafíos de adaptación para el desarrollo de la vitivinicultura, mientras que las provincias del sur y las regiones altas se verían beneficiadas por condiciones de cultivo favorables (Cabré y Nuñez, 2020).
Teslic et al. (2019) afirmaron que para el período 2011-2040, la región italiana de Emilia-Romagna seguirá siendo apta para la producción de uva en los escenarios RCP4.5 y RCP8.5, pero el período 2071-2100 probablemente será demasiado caluroso para mantener el rendimiento actual. y calidad de la uva.
Usando un enfoque similar, Cardell et al. (2019) encontraron que el aumento general proyectado para las temperaturas máximas en toda Europa podría conducir a la pérdida de viabilidad para la vinificación de algunas regiones en un futuro cercano si no se implementan medidas de adaptación. Esto se debe principalmente a que la planta está expuesta a temperaturas que exceden el umbral de maduración por períodos más prolongados (Cardell et al., 2019). Sin embargo, los países que actualmente tienen una temperatura del suelo más fría, como Alemania, el norte de Francia, Bélgica, el sur de Inglaterra y la República Checa, alcanzarían condiciones térmicas más favorables a mediados de siglo (Cardell et al., 2019).
Schultz (2016) ha argumentado que las suposiciones subyacentes sobre la idoneidad de ciertos conjuntos de clima, región y variedades para el cultivo de la vid son decisivas y se han basado en parte en el mal uso del índice de suma de temperatura, como HI. Aunque existe un límite inferior más claro de temperatura de crecimiento para cada variedad, el límite superior ha ido avanzando progresivamente en muchas regiones sin efectos negativos en la calidad del vino (Schultz, 2016).
Naulleau et al. (2021) notaron que las condiciones locales, como la capacidad de agua disponible del suelo y la disponibilidad de agua de riego, generalmente no se integran en los estudios de mapeo de idoneidad. Esto, junto con los conflictos de uso del suelo derivados de las políticas de conservación y la preferencia por otros cultivos, podrían convertirse en importantes limitantes para el establecimiento de nuevos territorios vitivinícolas (Naulleau et al., 2021). Otro aspecto que no se considera es la capacidad de la planta para aclimatarse a diferentes condiciones ambientales, que a su vez pueden ser epigenéticamente heredables (Marfil et al., 2019).
A modo de ejemplo, el calentamiento previsto para otras regiones más frías a finales de siglo, como es el caso de Escocia, todavía no hará plausible la producción de uva para vino debido al aumento concomitante de las precipitaciones en zonas ya húmedas (Dunn et al., 2019).

Altitud y fenología
La fenología es el estudio de los eventos periódicos (etapas de crecimiento y desarrollo) en los ciclos de vida biológicos y cómo estos son influenciados por el medio ambiente. Dicha información es necesaria para las decisiones sobre el sitio, la selección de cultivares y las prácticas culturales (Keller, 2020). Las principales etapas fenológicas de Vitis vinifera L. son: brotación, floración, fructificación, bayas del tamaño de un guisante, envero (inicio de la maduración) y cosecha (Coombe, 1995). Las etapas fenológicas están muy influenciadas por la temperatura, la amplitud térmica y la radiación solar, que en conjunto determinan la duración de las fenofases para una determinada variedad (Zapata et al., 2016).
Falcao et al. (2010) evaluaron la influencia de la altitud del viñedo (de 774 a 1.415 m s.n.m.) en la fenología y maduración de uvas cabernet sauvignon en Santa Catarina, Brasil (latitudes 26°–28° S). Informaron temperaturas medias del aire hasta 4°C más bajas en altitudes más altas, lo que retrasó el tiempo de cosecha, medido como concentración de sólidos solubles totales de bayas (°Brix), hasta 2 meses en comparación con altitudes más bajas. Muñiz et al. (2015) observaron la misma tendencia en regiones de gran altitud del sur de Brasil. El viñedo a mayor altitud (1.400 m s.n.m.) tuvo una temperatura media del aire hasta 4,4°C más baja, y recibió un 20% más de PAR y un 12% más de radiación total en el máximo diario en comparación con un viñedo a 950 m s.n.m. Esta variación en el sitio de cultivo afectó dramáticamente la duración de las fenofases, con un retraso de 50–56 días en las fechas de cosecha y 37 días desde la brotación hasta la plena floración de los cultivares merlot y cabernet Sauvignon.
En un estudio de terroir en Suiza, Rienth et al. (2020) analizaron cómo interactúan las condiciones pedoclimáticas con la fenología de la vid. Si bien hubo diferencias en la brotación y la floración impulsadas principalmente por la altitud (375-575 m s.n. m.), las variaciones climáticas de la cosecha en el estudio de 3 años no arrojaron resultados consistentes.
González Antivilo et al. (2017) demostraron marcadas diferencias en las amplitudes térmicas registradas entre dos regiones contrastantes de América (Mendoza, Argentina y Prosser, Estado de Washington, Estados Unidos). En Mendoza, tanto las temperaturas medias como las mínimas son más altas que en Prosser, lo que reduce el riesgo de daños por heladas durante el invierno. Sin embargo, la amplitud térmica considerablemente mayor a 766 m s.n.m. en Mendoza (hasta 10° más alto que Prosser a 117 m snm) significa que habrá días inusualmente calurosos durante el invierno, lo que puede causar deaclimatación al frío y lesiones por frío posteriores a fines del invierno (Gonzalez Antivilo et al., 2017) .
Gaiotti et al. (2018) evaluaron el efecto de temperaturas nocturnas más bajas en la calidad de la baya de uva corvina. Las plantas en macetas se transfirieron a una cámara frigorífica durante la noche comenzando antes del inicio de la coloración y continuaron hasta el envero completo, lo que retrasó el envero en aproximadamente 6 días.
El uso de modelos fenológicos para predecir la duración de las fenofases a partir de información de temperatura y radiación global se ha vuelto cada vez más frecuente (Ramos et al., 2018; Alikadic et al., 2019; Hall y Blackman, 2019; Leolini et al., 2020). Alikadic et al. (2019) describieron el desarrollo de la vid en la región vitivinícola de Trento, Italia, caracterizada por marcadas diferencias altitudinales, utilizando el modelo Fenovitis (Caffarra y Eccel, 2010). Estudiaron cinco variedades (pinot noir, sauvignon blanc, chardonnay, merlot, pinot gris) y evaluaron la correlación estadística entre la altitud (67-950 m s.n.m.) y el momento de la brotación, la cosecha y la duración del intervalo entre la brotación y la cosecha. El tiempo de cosecha estuvo notablemente influenciado por la altitud y las variedades, oscilando entre 6,27 y 7,16 días cada 100 m de elevación, mientras que el tiempo de brotación varió entre 0,85 y 2,88 días cada 100 m.
Según los experimentos realizados y la bibliografía actualizada al respecto, se puede proponer que, a escala global, un aumento en la altitud provoca un retraso de las fases fenológicas (figura 2A).
Los principales efectos se observan en los tiempos de cosecha seguidos por la brotación, lo que podría aumentar el riesgo de daños por granizo y heladas, respectivamente, al prolongar el período de estructuras vulnerables expuestas a la parte más fresca de la temporada de crecimiento.
Por otro lado, las fases fenológicas más largas pueden permitir que ciertos cultivares con ciclos de cultivo más cortos alcancen niveles enológicos superiores al ayudar a sincronizar la maduración de la uva con una acumulación suficiente de compuestos aromáticos y de color.
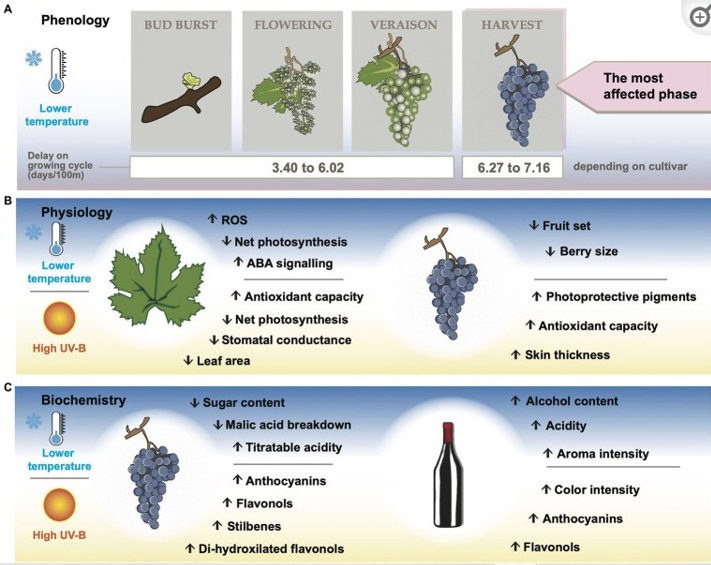
Altitud y fisiología
Los procesos fisiológicos clave afectados por las condiciones ambientales relacionadas con la altitud tienen lugar durante las etapas de elongación de los brotes y desarrollo de la baya (Pillet et al., 2015; Palliotti y Poni, 2016; Sawicki et al., 2016). La fisiología de la vid puede verse afectada (incluso dañando la planta), por factores estresantes o limitantes. Pueden incluir temperaturas excesivas en climas cálidos (especialmente en altitudes bajas), temperaturas bajas en climas templados/frescos y/o altitudes elevadas, y radiación UV-B solar extrema en altitudes muy elevadas (Körner, 2007; Figura 2B).
Las altas temperaturas pueden obstaculizar el crecimiento vegetativo y el desarrollo reproductivo de la vid, afectando el rendimiento y la calidad de la fruta. La fotosíntesis es uno de los procesos más sensibles al calor y, a menudo, se ve obstaculizada antes de que se manifiesten otros síntomas de estrés.
En general, las temperaturas superiores a 35 °C reducen la fotosíntesis en las hojas por el efecto combinado de la conductancia estomática restringida (Greer y Weedon, 2012), el centro de reacción del fotosistema II deteriorado y el estado de activación alterado de Rubisco (Luo et al., 2011). Por el contrario, las altitudes más altas pueden conllevar una mayor incidencia de días fríos y riesgo de heladas que se presentan no solo con temperaturas generalmente más bajas sino también con una mayor amplitud térmica (Körner, 2007).
Los brotes de vid de un año son particularmente sensibles a las lesiones a temperaturas inferiores a -2,5 °C, lo que provoca la pérdida de las yemas o flores primarias (Sawicki et al., 2016). Rooy et al. (2017) evaluaron los efectos del enfriamiento al disminuir gradualmente la temperatura del aire para tres cultivares de uva de mesa a 0°C y −4°C. Encontraron un aumento significativo en la fuga de electrolitos celulares y biomarcadores de estrés oxidativo (H2O2 y ácido tiobarbitúrico) en las hojas, supuestamente como resultado de la peroxidación lipídica y la lesión de las membranas (Rooy et al., 2017). Además, el enfriamiento puede reducir la conductancia estomática y la fotosíntesis neta (Sawicki et al., 2012; Król et al., 2015; Rooy et al., 2017), e incluso afectar el cuajado y la acumulación de carbono en las bayas (Sawicki et al., 2016).
También se ha observado un aumento en la concentración de prolina foliar y ABA durante la respuesta de tolerancia al frío en tres cultivares de vid (Rooy et al., 2017). Esta activación de la vía ABA puede conducir indirectamente a una mayor tolerancia al estrés abiótico debido a una mejora del sistema antioxidante de la planta (Berli y Bottini, 2013; Carvalho et al., 2015). Alonso et al. (2015) observaron este efecto al rociar ABA sobre hojas y bayas desde el envero en adelante, lo que promovió la acumulación de compuestos antioxidantes y antifúngicos.
Las temperaturas más frías en altitudes más altas también pueden extender los períodos de maduración de la baya, lo que favorece un recambio neto positivo de ácidos orgánicos, una mayor biosíntesis de flavonoles y antocianinas, lo que da como resultado una fruta más adecuada para la elaboración de vino tinto (Muniz et al., 2015; Martínez-Gil et al., 2018).
En las plantas, la luz actúa como fuente de energía para la fotosíntesis y como señal ambiental que provoca respuestas fotomorfogénicas. En altitudes más altas y durante el período de crecimiento de la vid, la PAR solar está mayormente por encima de los 700-900 μmol de fotones m−2 s−1, que es el punto de saturación de la vid. Las hojas expuestas a un exceso de luz suelen presentar una serie de mecanismos preventivos para evitar la fotoinhibición y el daño oxidativo.
Para disminuir la absorción de luz, las hojas pueden reducir el área de su lámina, ajustar su concentración de clorofila y acumular pigmentos fotoprotectores, como carotenoides y flavonoides, incluidas las antocianinas (Palliotti y Poni, 2016). Kok y Bahar (2015) describieron una disminución del 42% en el área foliar, junto con un aumento del 34% en la densidad estomática, en el cultivar Gamay desde los 405 m s.n.m. en comparación con las cultivadas a menor altura (180 m s.n.m.).
También se descubrió que la piel de la uva responde al exceso de radiación a través de cambios morfoquímicos. Un desnivel de 50 a 595 m s.n.m. causó una capa hipodérmica un 28% más gruesa en la piel de las bayas de Vranec (Nedelkovski et al., 2018). La hipodermis de las variedades tintas acumula antocianinas, que sirven como pigmentos fotoprotectores durante el desarrollo de la baya (Cadot et al., 2011; de Alencar Filho et al., 2016); y su acumulación se promueve a medida que aumenta la elevación (Berli et al., 2008).
Los viñedos de altura y más famosos de Mendoza, Argentina, se encuentran en Gualtallary a casi 1500 m s.n.m. (33°23′S, 69°15′W), con irradiancias UV-B que alcanzan hasta 40 μW cm−2 en los días de verano, mientras que a nivel del mar se mide alrededor de 15 μW cm−2 (Berli et al. , 2010).
El sistema de defensa de las hojas de vid frente a los rayos UV-B incluye la acumulación de pigmentos fenólicos fotoprotectores y la activación del mecanismo antioxidante enzimático y no enzimático, con la fitohormona ácido abscísico (ABA) actuando aguas abajo en la vía de señalización (Berli et al., 2010).
Los efectos de los rayos UV-B en la fisiología de la vid se evaluaron mediante experimentos de campo con exclusión solar de UV-B mediante láminas de plástico (desde la floración hasta la cosecha) durante diferentes temporadas de crecimiento (Berli et al., 2010).
Berly et al. (2013) encontraron que los efectos de los rayos UV-B en el crecimiento vegetativo son variables e interactúan con otras condiciones ambientales estacionales, como la temperatura del aire. En general, la radiación UV-B solar a gran altitud redujo la longitud de los brotes, la expansión de las hojas, la fotosíntesis y la conductancia estomática; y aumento del grosor de las hojas, pigmentos fotoprotectores, acumulación de prolina y capacidad antioxidante de las hojas (Berli et al., 2013; Martínez-Lüscher et al., 2013). Además, los altos niveles de UV-B redujeron el tamaño de la baya y el rendimiento de la fruta y mejoraron los compuestos fenólicos de la piel de la uva (Berli et al., 2011) y la capacidad antioxidante (Berli et al., 2015).
En un experimento similar con la variedad Graciano, Del Castillo Alonso et al. (2015) no encontraron diferencias entre las hojas sometidas a exclusión UV-B y los controles expuestos a UV-B. Esta falta de respuesta puede atribuirse a un diferencial UV-B insuficiente, un período de exclusión corto (en este experimento, el tratamiento comenzó en el envero), muestreo de hojas viejas (menos sensibles a los rayos UV-B), la supuesta capacidad de tolerancia de la variedad Graciano UV-B relativamente alta y/o una combinación de ellos. Esto puede implicar que la tolerancia UV-B depende del cultivo, o que la respuesta UV-B interactúa con otras variables ambientales que no se tienen en cuenta en estos experimentos.
Altitud y composición química de uvas y vinos
La mayoría de las características organolépticas del vino, a saber, color, aroma, sabores primarios y sensaciones en boca provienen de las características bioquímicas de la uva en la cosecha (Garrido y Borges, 2013; Wang y Spence, 2018). Los componentes químicos de la uva que son relevantes durante la vinificación son azúcares, compuestos fenólicos, ácidos orgánicos y compuestos orgánicos volátiles (Garrido y Borges, 2013; do Nascimento Silva et al., 2015; González-Barreiro et al., 2015). La Figura 2C proporciona una descripción general de cómo la temperatura más baja y la radiación UV-B más alta que se encuentran en los viñedos de gran altitud afectan la concentración de estos compuestos.
El contenido de azúcar de la uva está directamente relacionado con la síntesis, el transporte y la acumulación de metabolitos en los tejidos (Murcia et al., 2015; Mansour et al., 2022). Además, el azúcar de uva medido en °Brix con un refractómetro puede verse afectado indirectamente por los cambios en el contenido de agua y el tamaño de la baya.
Las bajas temperaturas en altitudes elevadas retrasan considerablemente la maduración de las bayas, además de que la menor fotosíntesis neta en las hojas expuestas a una mayor radiación UV-B provoca una menor acumulación de azúcar (Berli et al., 2011). Sin embargo, eso no necesariamente afecta la concentración de azúcar expresada en °Brix, ya que se observan mayores pérdidas por evaporación, aumentando la concentración de materia seca (Keller, 2010). Además, el tamaño de la baya tiende a disminuir con el aumento de la altitud como resultado de un mayor daño oxidativo, lo que finalmente contribuye a una mayor acumulación de azúcar por baya (Berli et al., 2011).
Según Robinson et al. (2011), en un estudio que compara la influencia del sitio, el manejo del dosel y la cepa de levadura, el principal factor que influye en la composición volátil y las características sensoriales de un vino es la ubicación del viñedo, que puede producir diferencias en hasta un 73 % de volátiles orgánicos del vino.
Además de las características específicas del sitio, como la composición del suelo y el aspecto del terreno, las principales variables ambientales que afectan la bioquímica de la uva son la temperatura y la exposición a la luz (Robinson et al., 2011; Garrido y Borges, 2013; González-Barreiro et al., 2015). Como se señaló anteriormente, estos varían de manera algo predecible con el aumento de la altitud (Körner, 2007), pero pueden variar de una manera más diferente según los cambios locales anuales en las condiciones ambientales.
Los compuestos fenólicos son uno de los tipos de metabolitos secundarios más abundantes e importantes en la uva y el vino, estando constituidos principalmente por ácidos hidroxicinámicos y flavonoides (Garrido y Borges, 2013). Hay tres grupos principales de compuestos flavonoides: proantocianidinas, antocianinas y flavonoles, y tienen funciones fisiológicas importantes durante el desarrollo de la baya, como actividad antioxidante, protección contra la radiación UV y defensa contra infecciones microbianas y fúngicas (Sun et al., 2017). Además, las antocianinas contribuyen principalmente a la pigmentación de los vinos, mientras que las proantocianidinas y los flavonoles son responsables de algunas de las principales propiedades sensoriales del vino, como la astringencia y la turbidez (Blancquaert et al., 2019).
Se ha reconocido que los factores ambientales, principalmente la luz y la temperatura, influyen en la composición fenólica de la uva y los vinos (Sun et al., 2017). Los estudios sobre la altitud de los viñedos han mostrado consistentemente una correlación positiva entre la altura y la producción de antocianinas y flavonoles (Berli et al., 2008; Li et al., 2011; Liang et al., 2014). En una serie de experimentos en cámaras de crecimiento dirigidos a determinar el rango de temperatura óptimo para la acumulación de antocianinas y flavonoles en las uvas merlot, Yan et al. (2020) encontraron que una amplitud térmica más alta, así como una temperatura nocturna más baja dieron como resultado los niveles más altos de flavonoles y antocianinas. Esto se relacionó con una expresión dos veces mayor de VviMybA, un factor de transcripción de antocianina central, durante el régimen de temperatura fría.
Particularmente, la mayor altitud tiende a promover antocianinas de tipo cianidina o no aciladas (Xing et al., 2016; Muñoz et al., 2021; Urvieta et al., 2021). Ciertamente, se ha señalado que la UV-B es el principal contribuyente a este efecto mediante la activación directa de la rama mediada por la flavonoide 3′-hidroxilasa (F3’H) de la ruta del fenilpropanoide en la piel de las bayas, que produce 3′,4′ -flavonoides hidroxilados como flavonoles tipo quercetina, antocianinas tipo cianidina, catequina y epicatequina (Berli et al., 2011; Martínez-Lüscher et al., 2014; Xing et al., 2016).
En un experimento de exclusión UV-B, vides de malbec que fueron expuestas a la luz solar UV-B a 1500 m s.n.m. tenía un 17,6 % más de polifenoles totales y un 28,5 % más de antocianinas totales en las pieles de las bayas en comparación con las del tratamiento con menos UV-B (Berli et al., 2008). En una configuración muy similar, las bayas de tempranillo a 371 m s.n. m. solo respondió levemente a la exclusión de UV-B, con una correlación positiva solo para la dosis de UV con los contenidos de flavonoles quercetina y kaempferol (Del-Castillo-Alonso et al., 2016). Esto puede atribuirse a la dosis bastante baja de UV-B a una altitud de 371 m s.n.m., más que a una suposición especulativa de que la variedad tempranillo se adapta bien a la UV-B.
La temperatura, por otro lado, parece seguir una correlación negativa con la concentración de flavonoides (Pastore et al., 2017). Esto probablemente se deba a que las temperaturas superiores a 40 °C inhiben la fenilalanina amoníaco liasa (PAL) y la estilbeno sintasa, enzimas clave de la ruta de los fenilpropanoides (Ferrandino y Lovisolo, 2014; Pastore et al., 2017). Mori et al. (2007) relacionaron este efecto inhibitorio de la temperatura alta tanto con una menor acumulación de ARNm de los genes de la vía de las antocianinas para ciertas variedades como con una degradación selectiva de las antocianinas no metiladas menos estables. Esto da como resultado un contenido de antocianina más bajo en general junto con un cambio en el tono favorecido por los derivados de malvidina altamente metilados. Como afirman Drappier et al. (2019), para una acumulación óptima de antocianinas, los racimos deben exponerse a temperaturas de 15 °C en la noche y 25 °C en el día durante la maduración. En consecuencia, Urvieta et al. (2021) encontraron una menor acumulación de antocianinas en uvas malbec y vinos cultivados en sitios de menor altitud de Mendoza (635 m snm), donde el número de días con temperaturas superiores a 33°C fue significativamente mayor en comparación con sitios de mayor altitud (1500 m snm).
Movahed et al. (2016) vincularon directamente este fenómeno tanto con la inhibición de la biosíntesis por regulación transcripcional como con la activación génica de las peroxidasas durante el estrés por calor, que median la degradación de las antocianinas. En un estudio de campo con un clima simulado de 2 a 3 °C más cálido que el control, de Rosas et al. (2017) encontraron una reducción significativa de antocianinas totales (28-41%) y una mayor proporción de antocianinas aciladas en pieles de uva malbec y bonarda. Esto se correlacionó con una menor expresión de genes de antocianina reguladores y estructurales.
Por el contrario, se sabe que las temperaturas más frías mejoran el color y la estabilidad de los vinos (González-Barreiro et al., 2015). Falcao et al. (2010) asociaron este fenómeno con el hecho de que los racimos permanecen más tiempo adheridos a la vid, por lo que tienen un envero prolongado para cosechar la fenofase para acumular compuestos coloreados en respuesta a la luz. Además, una baya más pequeña causada por las condiciones de gran altitud podría inclinar los compuestos polifenólicos a una concentración más alta cuando se analiza según la base de la baya (Berli et al., 2011). Variedades touriga nacional y touriga francesa cultivadas a 300–350 m s.n.m. contenían hasta un 59% más de antocianinas totales que las cultivadas a 100–150 m s.n.m., supuestamente debido a un diferencial de temperatura media de 5°C (Mateus et al., 2001a, 2002). Los autores argumentaron que las antocianinas, al interactuar con la también mayor cantidad de flavanoles, catequinas y procianidinas, mejoran la estabilización del color y la capacidad de envejecimiento en el vino elaborado con bayas cultivadas a mayor altitud (Mateus et al., 2001b).
Contrariamente a la mayoría de la literatura, Pajovic et al. (2014) encontraron para las variedades Vranac, Kratosija y cabernet sauvignon polifenoles totales más bajos y proantocianidinas tanto monoméricas como poliméricas en uvas cultivadas a 400 m s.n.m. en comparación con las cultivadas a 25 m s.n.m. Los autores propusieron que estos resultados se atribuyeron al hecho de que el cultivo se realizó en una región bastante fría y, por lo tanto, las bajas temperaturas fueron perjudiciales para el metabolismo secundario.
Los mayores contenidos de polifenoles en grandes altitudes también podrían estar relacionados con la activación de las respuestas de estrés abiótico dependientes de ABA a las bajas temperaturas, lo que en última instancia conduce a la acumulación de antioxidantes como el glutatión, los carotenoides y los flavonoides (Ferrandino y Lovisolo, 2014).
La relación positiva entre los factores ambientales relacionados con la altitud y el sistema de defensa antioxidante ha sido documentada para diferentes cultivares y regiones (Berli et al., 2015; Coklar et al., 2017), apuntando generalmente a un aumento de compuestos fenólicos con actividad antioxidante, sino también a una mayor actividad de las enzimas antioxidantes como respuesta al efecto combinado de mayor UV-B y menor temperatura que genera especies reactivas de oxígeno.
El pH del vino determina el equilibrio entre las diferentes estructuras de antocianinas. En condiciones muy ácidas, el catión flavilio (rojo) es la estructura principal de la antocianina, pero se reemplaza progresivamente por la base quinoidal (azul) a medida que aumenta el pH (Kontoudakis et al., 2011). Por lo tanto, la acidez titulable más alta y el pH más bajo que se encuentran en los mostos de ciertos cultivares cultivados a temperaturas más altas en comparación con los más bajos.
Fuentes: Frontiers in Plant Sciencia (Enoreports 182: «Climate Change Effects on Grapevine Physiology and Biochemistry: Benefits and Challenges of High Altitude as an Adaptation Strategy», por Arias, L. A.; Berli, F.; Fontana, A.; Bottini, R. y Piccoli, P.) y Ace Enología






